Sayer Ji
Jan 03, 2026

Read, comment, and share the X thread dedicated to this investigation.
Seasonal influenza is still interpreted through a framework that modern biology has already outgrown. Within that framework, symptoms such as fever, fatigue, inflammation, and withdrawal are treated as signs of pathological failure. Yet mounting evidence suggests these same responses may represent the body’s most powerful mechanisms of preservation, detoxification, and systemic reset—processes that keep the organism alive rather than threaten it.
Article-at-a-glance
Influenza-like illness can be read as a host-led program—a coordinated acute-phase reset shaped by immune signaling, metabolic cleanup, circadian timing, and environmental constraints—not merely a simplistic “invader enters, disease happens.”
Modern biology dissolves the hard boundary between “self” and “virus.” Influenza virions carry substantial host proteins and share features with endogenous vesicle traffic, tightening the conceptual link between virions, exosomes, and intercellular communication.¹²
Seasonality is not only about germs; it is also about missing ancestral inputs—UVB scarcity, disrupted light/dark timing, dry air/absolute humidity, and the loss of hot/cold cycling that historically trained human physiology.²¹²⁸
Interventions that restore “eustress cycles” (exercise, heat exposure/sauna, cold contrast, sleep/light regularity) often improve hard endpoints (respiratory illness burden, pneumonia risk, all-cause mortality) in ways that single-target strategies rarely match.⁸⁹¹¹
The investigative question modern flu season avoids

Every winter, a familiar story is broadcast with industrial confidence:
A virus is “circulating.”
You are “exposed.”
Exposure becomes an “infection.”
“Infection” becomes symptoms.
Symptoms become a problem to suppress—ideally prevented by a pharmaceutical product.
And yet, lived physiology keeps interrupting the narrative.
People who push themselves through brutally intense training sessions—hard HIIT intervals, hill sprints, long heat-soaked workouts—often report a transient constellation of “flu-like” symptoms: profuse sweating, nausea, chills, trembling, headache, malaise, a sense of impending doom. Then, if they recover well, the aftermath is the opposite of illness: clarity, appetite normalization, mood elevation, sleep depth, and the unmistakable feeling that something has been cleaned out and rebuilt.¹³¹⁴I describe surviving this “near death” exertion a regenerative wave, which for me can last for over 24 hours.
And this phenomenon you really have to live to truly understand, is a profound clue.

Because if “flu symptoms” can be produced—on demand—by voluntary, controlled stress (heat, exertion, metabolic intensity), then a serious scientific question follows:
What if seasonal influenza-like illness is not only a story about contagion, but also a story about the body being forced into a belated, involuntary restoration cycle—because the daily inputs that once maintained resilience (light, thermic contrast, movement variability, microbial contact) and provided the substrate for metabolic and immunological self-regulation — for countless millennia — have been flattened or completed disappeared by modern life?
This article, while questioning several foundational assumptions of classical virology, is not an exercise in denial. Rather, it represents an effort to move beyond a one-dimensional, pathogen-centric interpretation of disease—one that has proven insufficient to explain mounting biological complexity. That narrow framing has, perhaps too conveniently, supported both the continual expansion of vaccine schedules falsely and irresponsibly promoted as unequivocally “safe and effective,” and the recurring, seasonal cycle of media- and medicine-amplified fear, in which invisible particles are portrayed as the primary threats to human health, demanding passive vigilance rather than deeper inquiry into terrain, resilience, and causation.
What follows is an evidence-based “terrain-first” investigation built from three converging revolutions:
The holobiont revolution (microbiome + virome): we are ecosystems, not sealed individuals.¹⁶¹⁷¹⁸
The extracellular vesicle revolution (exosomes): biology moves information through virus-sized packages that look, behave, and sometimes blur into what we call “viruses.”²³⁴
The chronobiology + environmental physiology revolution: immunity is rhythmic, seasonal, and deeply dependent on light, temperature, humidity, sleep timing, and metabolic conditioning.²¹²⁸
Layer these together, and influenza becomes more than a villain. It becomes a pattern—and possibly, in many cases, a program for physiological resilience, without which perhaps our body would succumb to forces of degeneration and fragility.
Exhibit A: You are not an individual—at least not in the way medicine was trained to imagine

If the last century of medicine taught a cultural mantra—“You are you, and microbes are outsiders”—the microbiome shattered it.
A modern human is better described as a holobiont: a host plus a vast constellation of microbial life that participates in digestion, immune education, barrier maintenance, neurotransmitter metabolism, and even circadian signaling.¹⁷¹⁸²⁰
Even the numbers are psychologically destabilizing:
Updated quantitative estimates place bacterial cells in the body on the same order of magnitude as human cells—roughly comparable totals, not a trivial minority.¹⁸
The “self” you defend is genetically entangled with ancient viral history: a substantial portion of the human genome derives from sequences with similarity to retroviruses (endogenous retroviruses), representing a deep blur between “viral” and “human” at the level of inheritance.¹⁹
If your immune system is defending (and/or an extension of) an ecosystem—rather than a sterile fortress—then “infection” becomes less like invasion and more like ecological rebalancing: shifts in signaling, tolerance, barrier function, and microbial community dynamics.
This is not abstract philosophy. It changes the meaning of influenza-like disease.
Because in a holobiont frame, what matters is not merely “Did something enter?” but:
What state is the terrain in? (nutrient sufficiency, sleep, stress and toxicant load, metabolic flexibility)
What is the timing? (circadian gating of immune responses)²⁸
What environmental constraints are present? (UVB deprivation, indoor dryness, cold monotony, reduced thermic hormesis)²¹
What communication is occurring between cells? (extracellular vesicles, interferon programs, inflammatory myokines)³¹³
In other words, the body is not passively responding, but orchestrating a system-level process. Viruses, as obligate parasites, lack autonomous agency and cannot traverse biological space without being produced, packaged, and trafficked by host cells themselves. From this perspective, viral particles may frequently function as informational artifacts of host biology—signals embedded in cellular stress, repair, or adaptation pathways—rather than singular causal agents acting upon a helpless organism.
Exhibit B: The influenza “virus” is not purely “other”—it is built with “self”
Here is the single study that should permanently complicate simplistic “invader” metaphors.

A landmark proteomic analysis of influenza virions found that influenza particles maintain a conserved architecture across host/virus combinations—and that this “core” includes substantial quantities of host proteins, elaborated by abundant host-dependent features. The result: virions produced by mammalian and avian hosts show distinct protein compositions.¹
This is not a fringe claim. It is a direct experimental observation: influenza virions are materially hybrid—assembled through host machinery, incorporating host components.
Two implications matter for a newcomer trying to understand why an alternative view of viral particles can even exist:
“Virus” is not a neatly separable object like a bullet. It is more like a process—a replication/assembly logic that runs through host cellular pathways.
The boundary between virion and endogenous vesicle traffic is conceptually thin. If a particle’s membrane and protein cargo are strongly host-shaped—and if its size overlaps with endogenous communication vesicles—then confusion between “self-messaging” and “pathogen particle” is not insane; it is biologically plausible.²
This does not imply that influenza cannot be infectious. It requires, instead, a clarification of what we mean by infection. If infection is understood as the transfer of biologically meaningful information from one organism to another, then it is not inherently pathological. On the contrary, such exchanges are fundamental to communication, coordination, and adaptation across individuals, populations, and even species. Within this frame, the “infectious particle” occupies a continuum with endogenous signaling particles, and immune activation may be driven as much by patterns of distress signaling, barrier disruption, and inflammatory coordination as by simple molecular foreignness.
Once this distinction is recognized, influenza-like illness no longer appears as a biological war, but as a system-level orchestration.
Exhibit C: The exosome revolution—why a layperson is right to be confused
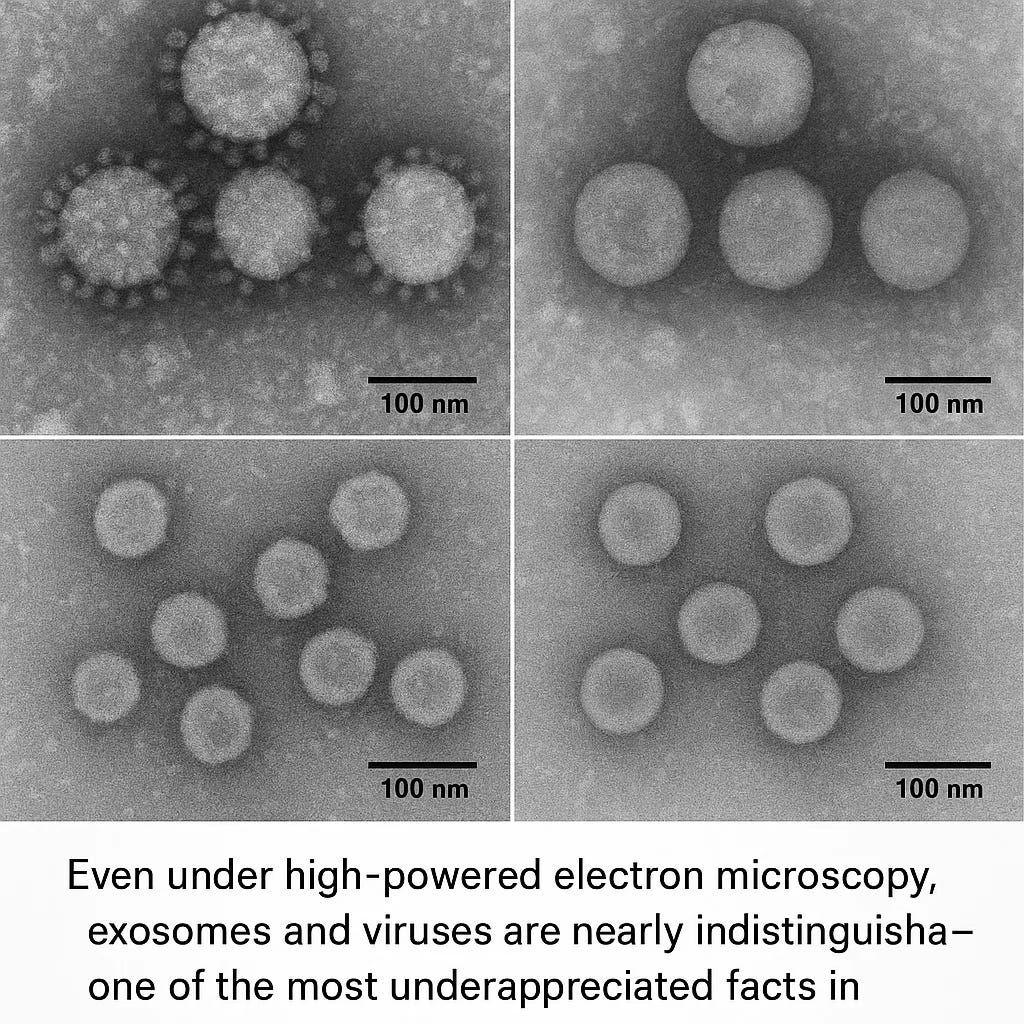
For most of medical history, only one kind of cell-to-cell communication felt “real”:
hormones in blood,
nerves firing,
cytokines as emergency flares.
But cells also communicate by shipping membrane-bound packages loaded with proteins, receptors, lipids, and genetic material (RNA). These are broadly called extracellular vesicles (EVs); a prominent subclass are exosomes.³
Exosomes matter here because they collide with virology at multiple levels:
They can be virus-sized.
They can carry nucleic acids and proteins that alter recipient cell behavior.³
Viruses can exploit EV biogenesis pathways; this is explicitly discussed in the literature (e.g., the “Trojan exosome hypothesis” for retroviruses).⁴
Scholarly reviews analyze the overlap directly: extracellular vesicles and viruses can be considered “close relatives” in formation pathways, composition, and functional mimicry.²
A person new to this terrain asks a fair question:
If cells naturally release virus-sized, genome-carrying vesicles—and viruses can use similar pathways—how clean is the practical distinction between “virus particle” and “communication particle,” especially in complex tissues under stress?
This is not a rhetorical loophole. It is one of the reasons modern virology increasingly discusses “viral infection” as an interplay of:
classical virions,
defective particles,
EVs carrying viral cargo,
EVs shaping immune responses,
and host-derived vesicles that can look “viral” by appearance or molecular signature.²⁶
Now add an additional observation: tissue injury can export its state through exosomes. In a striking experimental model of acetaminophen-induced liver injury, researchers investigated whether exogenous exosomes from injured mice could promote hepatotoxicity in recipients.⁵
Whether one interprets that finding as “toxicity messaging,” “damage-associated signaling,” or “exosomal programming,” the core point remains:
The body can transmit a disease-like state without requiring a living microbe to reproduce inside the recipient.
That mechanism—toxicant-induced state transfer—is central to why a “poisoned/not infected” reframe attracts attention. It offers a vocabulary for phenomena that look contagious, feel contagious, and cluster like contagion—yet may be substantially driven by shared exposures and shared signaling.
Go deeper here:
Reframing Viral Mechanisms: Exosomes, Toxicity, and the Xenogen Hypothesis
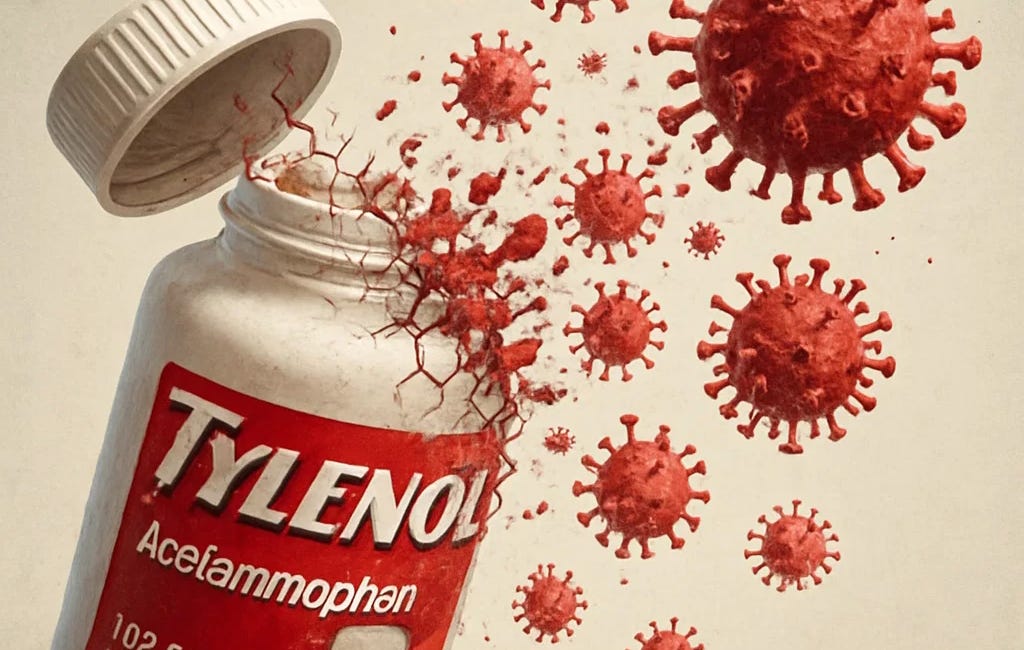
In a 2018 study published in Scientific Reports, researchers explored the effects of acetaminophen-induced liver injury in mice, with an initial focus on validating toxicological mechanisms of liver damage. However, their findings inadvertently cast light on a much deeper—and potentially paradigm-shifting—insight into the nature of so-called "viral" dis…
Exhibit D: “Poisoned Not Infected” as a model—what it claims, and what it attempts to explain
In the Poisoned Not Infected framework, the argument is not that pathogens are imaginary. The argument is that modern medicine often commits a category error:
It takes a complex systemic response (detoxification + repair + immune coordination + rhythmic resetting) and collapses it into a single word—infection—as if naming the event were explaining it.
Across the series, the proposed reframing is essentially this:
Many “viral syndromes” may be terrain crises: the body forced into emergency cleanup and adaptive signaling (even vaccines can induce such a crisis, effectively generating pseudoinfection and pseudocontagion)
Viral particles (or viral-like particles) may be part of information transfer in that response—sometimes as classical infection, sometimes as vesicle-mediated pattern propagation, sometimes as endogenous outputs that are misread as external attack.
The absence of daily ancestral inputs (thermic cycling, UVB, circadian stability, microbial richness) may create a pressure gradient that discharges seasonally, episodically, and dramatically—what we call “the flu.”³²³³
From a mainstream lens, some of these claims are speculative, some are provocative, and some are best read as hypothesis-generating rather than settled. But as an investigative project, the series is valuable because it insists on questions modern flu discourse prefers to skip:
Why do influenza-like epidemics correlate so tightly with seasonal environmental variables (humidity, temperature behavior, indoor crowding, UVB deprivation)?²¹
Why do symptoms often resemble a coordinated, metabolically expensive program—fever, anorexia, somnolence, myalgia—that looks designed, not accidental?⁶⁷
Why do interventions that restore ancestral stressors (movement intensity, heat exposure, thermic contrast) often correlate with profoundly reduced chronic disease and mortality—and sometimes reduced respiratory disease outcomes as well?⁸⁹¹¹
The “xenogen” inspiration in the model is, in effect, a modernized terrain theory that preserves an essential caveat:
Information transfer is real. Not all information is neutral or positive; some is dys-information (disease-promoting)
Essentially, biology is not only chemistry; it is messaging.
The dispute is not “does information move?” It is “how often do we mistake messaging and cleanup for invasion?”
Exhibit E: COVID as the stress test—why the argument expands beyond influenza

Influenza is the gateway; COVID is the accelerator.
The reason is simple: COVID pushed diagnostic technologies (especially PCR) into the center of public identity and fear, turning molecular detection into a social verdict.
Here is the important, mainstream-compatible clarification that most people never received:
PCR detects genetic material; it does not, by itself, prove the presence of live, replication-competent virus.
Clinical context matters. Viral culture, antigen tests, symptom timing, and epidemiology matter.
Once you accept that, two parallel truths can coexist without contradiction:
SARS‑CoV‑2 is supported as a real infectious agent by multiple lines of evidence (not just PCR).
PCR positivity can sometimes reflect residual fragments or low-level signals that do not map neatly onto “you are infected in the way you imagine infection.”
The Poisoned Not Infected series uses COVID as a case study to argue that exosomes, fragments, and host-derived nucleic acids may inflate the conceptual certainty people place on molecular signals—and that this can distort both policy and personal fear.³²³³
The Poisoned Not Infected Framework: A Four-Part Reorientation of Disease

Part I: Poisoned, Not Infected—Why Your Body’s Healing Response Looks Like Disease
This installment lays the conceptual groundwork: “viral disease” can be reframed as a healing response that is routinely misinterpreted as pathology, with heavy emphasis on extracellular vesicles, endogenous messaging, and the interpretive limits of single-test epistemology.³²
Part II: Poisoned Not Infected—Reinterpreting Viral Disease Through Chemical Toxicity
This section extends the argument into environmental medicine: clustered illness can emerge from shared toxicant exposure and shared physiologic stressors—producing outbreaks that resemble contagion.³³
Part III (adjacent): When the Vaccinated Body Becomes the Broadcast Tower—The Shedding Paradox
This piece explores a controversial hypothesis: whether vaccine-driven expression of antigens could, via vesicles or other mechanisms, contribute to symptom patterns in ways that resemble transmission.³⁵
A scientifically cautious reader should treat this as speculative: it proposes a mechanism that is not established as a dominant driver of population illness patterns. But it is included here because it illustrates the series’ consistent theme: the body exports states through biology that is not always captured by the word “infection.”
Part IV: The Long COVID essay: Unmasking the “Long COVID” Cover Story for Vaccine Injury
This essay argues that long COVID narratives may obscure vaccine-related harms.³⁴
Whatever one’s position, the key investigative value is that it forces differentiation between:
post-infectious syndromes,
post-vaccination adverse events,
and broader dysautonomia/inflammatory states that can be triggered by multiple upstream stressors.
A responsible synthesis acknowledges that long COVID is observed in unvaccinated people as well, and that many studies suggest vaccination reduces long COVID risk on average—while still leaving room for legitimate scientific work on adverse events and heterogeneity of outcomes.
The deeper point for influenza: once you see how “one label” can hide multiple mechanisms, you stop assuming the seasonal flu is a single mechanism event.
Exhibit F: Seasonality is a physiology problem, not only a pathogen problem

If contagion were the primary driver of seasonality, we would expect something closer to a constant baseline with random spikes. Instead, influenza behaves like a system constrained by environmental variables—particularly those that modulate airway biology and immune timing.
1) Absolute humidity and the physics of winter outbreaks
Research shows that absolute humidity strongly constrains influenza survival/transmission and seasonality dynamics.²¹
This is not “terrain theory.” This is mainstream epidemiology: cold-season air is typically drier, and indoor heating amplifies dryness, changing airway surface properties and viral particle/exosome stability.
2) UVB scarcity, vitamin D signaling, and “spectral deficiency”
The UVB question is not cosmetic. UVB is a biologically rare band of sunlight with disproportionate effects on endocrine and immune signaling.
The argument in the UVB-focused work you referenced is that modern humans are spectrally deprived, and that the seasonal loss of UVB is one of the deepest upstream drivers of winter immune instability.³¹
You also asked to include your presentation on this theme: The Most Powerful “Supplement” on Earth—a framing meant to shock people out of supplement fetishism by pointing to sunlight as a primary input.³⁰
Whether one fully agrees or not, the core point is difficult to dispute:
Light is not merely illumination; it is biological instruction.
Circadian entrainment, immune cell trafficking, hormone secretion, and inflammatory thresholds are all time-structured.²⁸
3) Circadian immunity: the immune system is a clockwork
A mature immune system is not “always on.” It is rhythmically gated—a property so robust that the field now discusses chrono-immunology and time-dependent immune interventions.²⁸
If winter disrupts sleep timing, morning light exposure, and daily activity rhythms—while adding psychological stress and reducing outdoor movement—then the immune system is being hit from multiple angles, even before any virion appears.
4) The missing thermic rhythm: we removed heat/cold cycling from daily life
Humans evolved with profound temperature variability: cold mornings, heat from exertion, sweat as routine, and periodic deep heat exposure through environment and labor.

Modernity flattened that into:
climate control,
sedentary monotony,
a narrow thermal band,
and fewer hormetic stress signals.
The terrain argument is straightforward:
If the body expects rhythmic thermic “training signals” and does not receive them, it may eventually deliver them to itself—sometimes violently—through involuntary fever programs and inflammatory resets.
Mainstream physiology already grants part of this: fever-range temperature can inhibit viral replication and influence immune function, even though fever can be costly and sometimes harmful in severe disease.⁶⁷
The terrain model extends this interpretation further: fever is not merely a defensive reaction to an external invader, but a deliberate tool of systemic recalibration. As Dr. Thomas Cowan describes in The Contagion Myth, elevated body temperature alters the physical state of intracellular and interfacial water—particularly the gel-like, structured water often described as the fourth phase—rendering accumulated toxicants and damaged nucleic acids (mistaken for ‘viral’ via PCR) more soluble and, therefore, more readily excretable. From this perspective, fever enhances the body’s capacity to mobilize and eliminate substances whose persistence could otherwise contribute to severe, even fatal, illness. In such a context, fever and influenza-like symptom clusters may function not as signs of biological failure, but as potentially life-preserving processes.
I explore this theme further in REGENERATE, where I describe the so-called sickness syndrome as one of the few remaining mechanisms by which modern humans are compelled to rest, withdraw metabolic resources from outward productivity, and allocate sufficient time and energy toward detoxification, repair, and regeneration. In a culture structurally resistant to rest and recovery, illness may represent not an aberration, but the body’s last enforced opportunity to heal.
Exhibit G: Eustress as “prehabilitation”—why heat, exercise, and contrast matter

We now return to the opening clue: why hard training can feel like the flu—and why it can also prevent the flu-like crash from becoming catastrophic.
1) Intense exercise produces “sickness signals”—and then resolves them
High-intensity exercise evokes a transient physiological state that closely resembles acute illness. During maximal or near-maximal exertion, the body generates cytokine signaling, stress hormones, metabolic byproducts, and short-lived immune perturbations. In doing so, the organism is forced to practice key adaptive processes, including:
inflammation onset and resolution, rather than chronic inflammation
oxidative stress handling, including upregulation of endogenous antioxidant systems
mitochondrial throughput and efficiency under load
lactate and hydrogen ion (H⁺) tolerance, with rapid shifts in acid–base balance
recovery-driven rebuilding, including tissue repair and metabolic recalibration
Skeletal muscle is now understood to function as a powerful endocrine organ, releasing myokines—most notably interleukin-6 (IL-6)—that participate in systemic metabolic coordination and immune regulation. Importantly, IL-6 released during exercise behaves differently from IL-6 released during chronic inflammation, acting instead as a signal that helps initiate resolution and repair rather than pathology.¹³
The nausea, dizziness, and malaise that often accompany all-out efforts are not imagined side effects. Lactic acidosis and rapid metabolic flux have been shown to correlate strongly with sprint-mediated nausea and flu-like sensations in controlled studies.¹⁴
When heat exposure (such as sauna) or cold immersion is layered onto intense exercise, the adaptive signal deepens further. Heat induces heat-shock proteins (HSPs)—including HSP70 and HSP90—which act as molecular chaperones that stabilize damaged proteins, facilitate repair and autophagy, and improve cellular stress tolerance. Cold exposure, in turn, induces cold-shock proteins (such as RBM3), which support RNA stability, mitochondrial efficiency, and cellular resilience under metabolic stress. Together, these stress proteins encode a kind of molecular memory of survival, teaching cells how to recover from challenge rather than collapse under it.
In essence:
A hard HIIT session can mimic “illness” because it is a controlled rehearsal of crisis physiology.
Done intelligently—with sufficient recovery—this rehearsal trains the organism not only to enter a high-stress state, but to exit it cleanly and efficiently. In this way, regular exposure to eustress may reduce the need for a more chaotic, involuntary, system-wide reset—what we recognize as influenza-like illness—to restore capacities that are otherwise lost in a modern lifestyle stripped of ancestral stressors. This interpretation is consistent with available biomedical literature linking physical conditioning, metabolic flexibility, and thermic hormesis with reduced illness burden.

2) But there is a boundary: the J-curve and the cost of excess
The literature also cautions that unusually heavy acute or chronic exercise—especially when paired with inadequate recovery—can increase the risk of upper respiratory tract infections (URTIs) in some contexts, a relationship often described as a **“J-shaped curve.”**¹²
This nuance strengthens, rather than weakens, the terrain model. The goal is not relentless stress, but eustress.
Eustress is a pulse:
intensity → recovery → adaptation.
When stress is continuous, unresolved, or layered onto sleep deprivation and nutritional deficit, it becomes distress, and resilience erodes. Influenza-like illness, in this frame, may represent the body stepping in to enforce a reset that voluntary practices failed to provide.
3) Exercise training can reduce respiratory infection burden
Randomized and controlled evidence suggests that structured training interventions—including exercise programs—can reduce aspects of acute respiratory infection incidence, duration, and severity, though effects vary and many studies are underpowered.¹¹
The direction of effect is nevertheless consistent with a terrain-based interpretation:
more resilient physiology, fewer severe illness days, and a reduced likelihood that a respiratory challenge escalates into a full systemic crisis.
4) Sauna and heat therapy: mortality data and respiratory associations
Heat exposure is not folklore—it carries serious observational outcome signals.
Frequent sauna bathing is associated with substantially reduced risks of fatal cardiovascular events and all-cause mortality in large Finnish cohorts.⁸
Separate prospective analyses link regular sauna use with reduced risk of pneumonia and other respiratory diseases.⁹¹⁰
Is this proof that sauna “prevents influenza”? No. But it is strong evidence that heat exposure functions as a broad resilience practice, one that appears to lower the risk of the outcomes that matter most—severe illness and death—rather than merely altering test positivity or symptom counts.
This directly supports your rhetorical challenge:
If a strategy improves mortality and systemic resilience, it deserves a central place in prevention discourse—especially when compared with interventions whose absolute benefits may be modest in low-risk populations.
5) Colostrum: a supportive tool, not a worldview
Bovine colostrum has clinical trial and meta-analytic evidence suggesting reductions in upper respiratory symptoms in certain populations, particularly athletes under heavy training loads.¹⁵
This can reasonably be interpreted as immune support, barrier reinforcement, or mucosal resilience.
But the deeper terrain point remains: colostrum is not the paradigm—it is a lever.
The paradigm shift is to stop treating “natural prevention” as a gentler extension of germ warfare, and instead recognize these tools as means of restoring terrain, rhythm, and adaptive capacity—thereby reducing the likelihood that the body must resort to dramatic, involuntary illness to recalibrate itself.
Beyond the Flu Shot: How Colostrum Challenges the 'Holy Cow' of Vaccine Dogma

Read, comment and share the X thread dedicated to this article here: https://x.com/sayerjigmi/status/1970328177579823131
Exhibit H: What about vaccines—how to discuss them without propaganda
Any serious examination of influenza prevention must confront a question that is typically avoided rather than answered: why assume the risk of an influenza vaccine at all if non-pharmaceutical strategies exist—and if influenza vaccination has never demonstrated a profound, reliable reduction in mortality?
An honest investigative analysis must begin with a methodological reality that is rarely stated plainly:
Claims of benefit are only as strong as the integrity of the systems that generate them.
Influenza vaccine research operates within an ecosystem shaped by industry funding, regulatory capture, publication bias, endpoint selection, and structural incentives that consistently favor positive interpretation. Under such conditions, neutrality cannot be presumed.
Several features of the evidence base are therefore not reassuring, but disqualifying for strong claims:
Vaccine effectiveness estimates fluctuate widely by season, strain selection, population, and outcome measured—suggesting instability rather than robust protection.
Systematic reviews in healthy adults repeatedly find only modest or negligible effects on influenza-like illness, while emphasizing the absence of convincing evidence for reductions in serious complications, hospitalization, or all-cause mortality.
Observational studies reporting benefits often rely on designs highly vulnerable to bias, including healthy-user effects, confounding by healthcare-seeking behavior, differential testing, and selective outcome reporting.
Counterintuitive or negative findings regularly emerge in independent analyses—particularly in working-age adults—yet are frequently reframed, dismissed, or buried rather than prompting structural reevaluation.
To accept influenza vaccination as “beneficial” in any meaningful sense would require assuming that these distortions are either minimal or accidental—an assumption that is increasingly implausible given the political, financial, and institutional forces involved.
From a terrain-first perspective, the deeper issue is not whether influenza vaccines provide any effect under narrow conditions, but whether they meaningfully address causation.
They do not.
Vaccination targets a hypothesized agent, not the systemic conditions that determine whether illness emerges, escalates, or resolves. As such, it cannot plausibly substitute for interventions that demonstrably improve resilience across diseases and decades.
A terrain-centered public message therefore does not need to posture as “anti-vaccine” to be truthful. It can simply state:
Health should not be outsourced to a recurring pharmaceutical intervention whose benefits are uncertain, seasonally contingent, and unsupported by mortality data.
Risk stratification does not rescue a flawed model; it merely narrows its application without addressing its conceptual failures.
The most powerful and controllable determinants of health remain terrain drivers: sleep timing, circadian light exposure, movement intensity and recovery, metabolic flexibility, thermic hormesis, micronutrient sufficiency, toxicant burden, and stress regulation.
In short, influenza vaccination persists not because it has solved the problem of seasonal illness, but because the framework that would replace it has not yet been permitted to fully emerge.
This article does not argue that influenza vaccines “should not exist.”
It argues that they cannot be honestly positioned as a cornerstone of prevention—and that continuing to frame them as such delays the far more consequential work of restoring biological coherence in a population chronically deprived of the conditions required for health.
The terrain conversation is not an adjunct.
It is the conversation that was postponed.
An Inconvenient Arithmetic the Policy Narrative Rarely Acknowledges
One of the most embarrassing yet underdiscussed facts in the influenza literature appears not in fringe critiques, but in the plain language of the Cochrane reviews themselves.

Cochrane acknowledges that over 200 distinct viruses are believed to cause influenza-like illness (ILI), all producing essentially indistinguishable clinical symptoms—fever, headache, myalgia, cough, malaise, and rhinorrhea. Without laboratory testing, clinicians cannot reliably differentiate among these viruses, which typically cause self-limited illness lasting days and only rarely progress to severe disease.
Against this backdrop, a critical limitation emerges:
At best, seasonal influenza vaccines target only influenza A and B—together representing approximately 5% of all circulating viruses responsible for ILI.
Even before questions of efficacy, safety, strain mismatch, or study integrity are raised, this creates a hard probabilistic ceiling on what such vaccines could theoretically accomplish.
Seasonal influenza vaccines are typically trivalent or quadrivalent, formulated months in advance to include two influenza A strains and one or two influenza B strains projected to circulate in the upcoming season. The formulation assumes not only that these strains will dominate, but that they will do so sufficiently to justify mass intervention—an assumption that frequently fails in practice.
From a purely statistical standpoint, the maximum theoretical benefit of a trivalent influenza vaccine—assuming perfect strain match, perfect uptake, perfect immune response, and zero adverse effects—would be confined to a small fraction of all influenza-like illness, and an even smaller fraction of total respiratory illness burden.
In real-world conditions, where:
strain prediction is imperfect,
immune responses vary widely,
circulating pathogens shift dynamically, and
influenza A and B often represent a minority of ILI cases,
the expected benefit collapses further.
This arithmetic exposes a foundational problem: influenza vaccination is routinely promoted as if it addresses “the flu,” when in fact it targets a narrow subset of viruses that cause a minority of flu-like illness. The public is encouraged to interpret any winter respiratory illness as evidence of vaccine “failure” rather than evidence that the intervention was never designed to address most causes of the syndrome in the first place.
From a terrain-first perspective, this is not a trivial oversight—it is a category error.
It shifts attention away from the shared physiological response (the syndrome) and toward a narrow molecular target, reinforcing the illusion that seasonal illness is primarily a problem of pathogen selection rather than host state, environmental mismatch, and systemic resilience.
When an intervention can, at best, apply to a small slice of the causal landscape—and when even that slice is subject to substantial uncertainty—it cannot reasonably be positioned as a cornerstone of prevention. The continued elevation of influenza vaccination to that status therefore reflects not biological necessity, but institutional inertia and narrative convenience.
In contrast, interventions that improve terrain—sleep, light timing, metabolic flexibility, movement intensity with recovery, thermic hormesis, micronutrient sufficiency—apply across all causes of ILI, regardless of viral taxonomy.
That distinction matters. Because biology responds to conditions, not labels.
A synthesis model: influenza as “trigger + terrain + timing + signal”

If you want a paradigm-shifting frame that preserves complexity without drifting into denial, it looks like this:
1) Trigger
A trigger can be a virus, a co-infection, a pollutant exposure, sleep debt, metabolic crash, or a cluster of winter constraints.
2) Terrain
The baseline state—nutrients, mitochondrial function, stress load, microbiome/virome balance, barrier integrity—determines whether the trigger becomes a minor event or a systemic crisis.
3) Timing
Circadian and seasonal timing gates immune responses.²⁸
Winter shifts timing, light exposure, and hormonal rhythms.
4) Signal
Cells communicate states. Exosomes/EVs transmit instructions and distress patterns.³²
Virions themselves can be interpreted as part of a broader continuum of biological messaging objects.¹²
In this model, “flu” is not always “the pathogen.” It is often the system-level output of a constrained organism forced to re-establish coherence.
Practical terrain restoration without making “flu” the enemy

This section is intentionally conservative: not medical advice, not a protocol, not a replacement for clinical care.
It is simply the terrain logic applied.
The goal: reintroduce the missing pulses
Daily light timing: morning outdoor light; consistent sleep/wake.²⁸³¹
Thermic hormesis: sauna or hot bathing, and (where safe) cold contrast—**a pulse, not punishment.**⁸⁹
Intensity + recovery: HIIT or hard efforts 1–3x/week depending on training status, paired with genuine recovery (sleep, hydration, nutrition).¹¹¹²
Microbiome literacy: food diversity, fiber, fermented foods, reduced ultra-processed load—because you are managing an ecosystem, not a machine.¹⁷¹⁶
Respiratory-season realism: humidity management indoors; ventilation; outdoor time; stress reduction.²¹
The terrain reframe is not “never get sick.” It is:
Make the reset smaller, cleaner, and less traumatic—so your body does not need to stage a dramatic seasonal intervention to regain balance. Or, better yet, live a lifestyle that is truly preventative. There are people who never get the flu, or only once every decade or so. Why is that? What lifestyle are they living? What can you do to implement healthy eating, mind set, exercise, and detoxification/rejuvenative regimens that make the need for ‘seasonal’ cold and flu obsolete?
Conclusion: the end of “germ theater,” the beginning of ecological medicine

A century ago, it was psychologically and politically useful to conceptualize disease as an external enemy. That framing helped mobilize sanitation infrastructure, antibiotics, acute emergency medicine, and certain targeted interventions—some of which undoubtedly reduced mortality from infectious crises that once overwhelmed populations. Yet the same adversarial model, when elevated to an unquestioned doctrine, also produced unintended consequences. In particular, the steady expansion of vaccine schedules—routinely promoted as uniformly “safe and effective” despite limited long-term, systems-level evaluation—has coincided with a profound epidemiological shift: the decline of many naturally self-limiting childhood illnesses (e.g. chicken pox and measles) alongside a marked rise in chronic neurological, immunological, and inflammatory conditions. Rather than eliminating disease, this tradeoff appears to have reorganized it, exchanging acute, time-limited challenges of development for lifelong disorders of immune regulation, neurodevelopment, and metabolic resilience—outcomes borne disproportionately by those who were encouraged to comply without being invited to question first principles.
But influenza season reveals the cost of staying trapped in that metaphor of foreign invasion by invisible particles.
Because the deeper truth is uncomfortable:
You are a holobiont.
Your genome contains viral history.
Your cells ship information in virus-sized packages.
Your immunity is rhythmic, seasonal, and environmentally trained.
And the symptoms you fear may be part of an ancient intelligence trying to restore coherence when daily inputs fail.
Influenza, in this view, is not a moral failure. It is a signal.
And the highest form of prevention is not panic. It is restoration of more ancient ways of self-care and resilience.

Learn more about natural approaches to influenza by using the GreenMedInfo.com database on the subject below.


Endnotes
Edward C. Hutchinson et al., “Conserved and Host-Specific Features of Influenza Virion Architecture,” Nature Communications 5 (2014): 4816.
Esther Nolte-’t Hoen et al., “Extracellular Vesicles and Viruses: Are They Close Relatives?” Proceedings of the National Academy of Sciences 113, no. 33 (2016): 9155–9161.
Clotilde Théry, Michal Ostrowski, and Elodie Segura, “Membrane Vesicles as Conveyors of Immune Responses,” Nature Reviews Immunology 9, no. 8 (2009): 581–593.
Stephen J. Gould, Melissa Booth, and Kim J. Hildreth, “The Trojan Exosome Hypothesis,” PNAS 100, no. 19 (2003): 10592–10597.
Young-Eun Cho et al., “Exogenous Exosomes from Mice with Acetaminophen-Induced Liver Injury Promote Toxicity in the Recipient Hepatocytes and Mice,” Scientific Reports 8 (2018): 15015.
Masamichi Yamaya et al., “Effects of High Temperature on Pandemic and Seasonal Influenza Virus Replication and Infection-Induced Damage in Primary Cultures of Human Tracheal Epithelial Cells,” PLoS ONE (2019).
L. Belon et al., “Effect of a Fever in Viral Infections,” Frontiers in Bioscience (2021).
Timo Laukkanen et al., “Association Between Sauna Bathing and Fatal Cardiovascular and All-Cause Mortality Events,” JAMA Internal Medicine 175, no. 4 (2015): 542–548.
Setor K. Kunutsor et al., “Frequent Sauna Bathing May Reduce the Risk of Pneumonia in Middle-Aged Caucasian Men,” Respiratory Medicine 132 (2017): 161–163.
Setor K. Kunutsor et al., “Sauna Bathing Reduces the Risk of Respiratory Diseases: A Long-Term Prospective Cohort Study,” European Journal of Epidemiology (2017).
Bruce Barrett et al., “Meditation or Exercise for Preventing Acute Respiratory Infection: A Randomized Controlled Trial,” Annals of Family Medicine 10, no. 4 (2012): 337–346.
David C. Nieman, “Exercise, Upper Respiratory Tract Infection, and the Immune System,” Medicine & Science in Sports & Exercise (1994).
Bente K. Pedersen and Mark A. Febbraio, “Muscle as an Endocrine Organ: Focus on Muscle-Derived Interleukin-6,” Physiological Reviews (2008).
R. J. Merrells et al., “Role of Lactic Acidosis as a Mediator of Sprint-Mediated Nausea,” Physiological Reports(2019).
A. W. Jones et al., “Bovine Colostrum Supplementation and Upper Respiratory Symptoms During Exercise Training: A Systematic Review and Meta-Analysis of Randomised Controlled Trials,” BMC Sports Science, Medicine and Rehabilitation 8 (2016).
Guohong Liang and Frederic D. Bushman, “The Human Virome: Assembly, Composition and Host Interactions,” Nature Reviews Microbiology (2021).
Human Microbiome Project Consortium, “A Framework for Human Microbiome Research,” Nature (2012).
Ron Sender, Shai Fuchs, and Ron Milo, “Revised Estimates for the Number of Human and Bacteria Cells in the Body,” PLOS Biology (2016).
David J. Griffiths, “Endogenous Retroviruses in the Human Genome Sequence,” Genome Biology (2001).
Eugene Rosenberg and Ilana Zilber-Rosenberg, “The Hologenome Concept of Evolution After 10 Years,” Microbiome (2018).
Jeffrey Shaman and Melvin Kohn, “Absolute Humidity Modulates Influenza Survival, Transmission, and Seasonality,” PNAS 106, no. 9 (2009): 3243–3248.
Milton J. Rosenau, “Experiments to Determine Mode of Spread of Influenza,” JAMA (1919); see also J. M. Eyler, “The State of Science, Microbiology, and Vaccines Circa 1918,” Public Health Reports (2010).
A. C. Sherman et al., “The Future of Flu: A Review of the Human Challenge Model,” Vaccines (2019).
Vittorio Demicheli et al., “Vaccines for Preventing Influenza in Healthy Adults,” Cochrane Database of Systematic Reviews (latest update). (Cochrane Library)
N. Rezaei-Tavabe et al., “The Effect of Monovalent Influenza Vaccine on the Risk of Hospitalization and All-Cause Mortality: A Meta-Analysis,” (2023).
N. K. Shrestha et al., “Effectiveness of the Influenza Vaccine During the 2024–2025 Respiratory Viral Season Among Working-Age Adults,” medRxiv (2025).
Rhonda Patrick and Jari A. Laukkanen (review), “Sauna Use as a Lifestyle Practice to Extend Healthspan,” American Journal of Physiology / review literature (2021).
Christoph Scheiermann, James Gibbs, and Arne N. Ince (review), “Clocking In to Immunity,” Nature Reviews Immunology (2018).
John J. Cannell, “On the Epidemiology of Influenza,” (2008).
Sayer Ji, “The Most Powerful ‘Supplement’ on Earth: Why Full-Spectrum Sunlight—and Especially UVB—May Be the Missing Frequency in Modern Health,” Substack (2025).
Sayer Ji, “Why UVB—The Rare Slice of Sunlight That Programs Human Biology—May Be the Root Cause You’ve Never Considered,” PDF upload.
Sayer Ji, “Poisoned, Not Infected: Why Your Body’s Healing Response Looks Like Disease,” PDF upload.
Sayer Ji, “Poisoned Not Infected: Reinterpreting Viral Disease Through Chemical Toxicity,” PDF upload.
Sayer Ji, “Unmasking the ‘Long COVID’ Cover Story for Vaccine Injury,” PDF upload.
Sayer Ji, “When the Vaccinated Body Becomes the Broadcast Tower: The Shedding Paradox,”
Sayer Ji, “How the Microbiome Undermines the Ego, Vaccine Policy, and Patriarchy,”
GreenMedInfo, “Sweating Out Heart Disease: The Cardiovascular Case for Infrared,”
No hay comentarios:
Publicar un comentario