Sayer Ji
Jun 09, 2026

For most of my adult life I treated energy the way most people do, as something to summon on demand. A second cup when the morning thinned out. Something sweet at three in the afternoon. I built a lot of work on borrowed energy before I understood that borrowing was exactly what I was doing, and that every loan carried interest my cells were quietly paying. This is the explanation I wish someone had handed me twenty years ago.
Why “Energy” Is the Most Misunderstood Word in Modern Health
When most people say they want more energy, what they actually mean is I want to feel awake, focused, and unburdened for more hours of the day — a perfectly reasonable thing to want, but the way modern culture has tried to deliver it, through coffee, tea, energy drinks, sugar, alcohol, and nicotine, is built on a category error.
The substances in that list do not generate energy. They borrow it. They block the chemical signals that tell your brain it’s tired (Ultrahuman blog on caffeine and adenosine). They flood the bloodstream with fuel your body wasn’t asking for (Mayo Clinic on reactive hypoglycemia). They occupy receptors that release dopamine without doing the cellular work that dopamine release normally signals (Biochemical Pharmacology on nicotine receptor upregulation). In every case, the feeling of energy arrives while the cellular machinery that produces energy either stays the same, is put under added strain, or becomes less efficient over time. The bill comes due as a crash, a tolerance escalation, or a pattern of ‘needing more to feel the same. (Gastroenterology — alcohol and mitochondrial dysfunction).
Physiologically, the feeling of energy arrives, while the actual cellular machinery that produces energy either stays the same, runs into debt, or gets damaged. The bill comes due as a crash, a withdrawal, a tolerance escalation, or — over years — measurable mitochondrial decline.
MethylateBLUE™ was built around a completely different premise. Instead of stimulating the perception of energy, it feeds the cellular systems that produce energy: the mitochondria, the methylation cycle, the NAD+ pool, the antioxidant enzymes, and the trace-mineral cofactors every redox reaction in the body depends on. The lift is slower to arrive and much slower to wear off, because it is the real thing, not a signal trick.
This article walks through every ingredient in the formula and explains, mechanism by mechanism, why this approach yields sustained, focused energy that the stimulant model structurally cannot.
MethylateBLUE™ is a dietary supplement intended to support normal mitochondrial and metabolic function. It is not intended to diagnose, treat, cure, or prevent any disease. Individual experiences vary.
The Cellular Logic of Real Energy
Before the ingredients make sense, the biology has to. And the biology is older than you are.
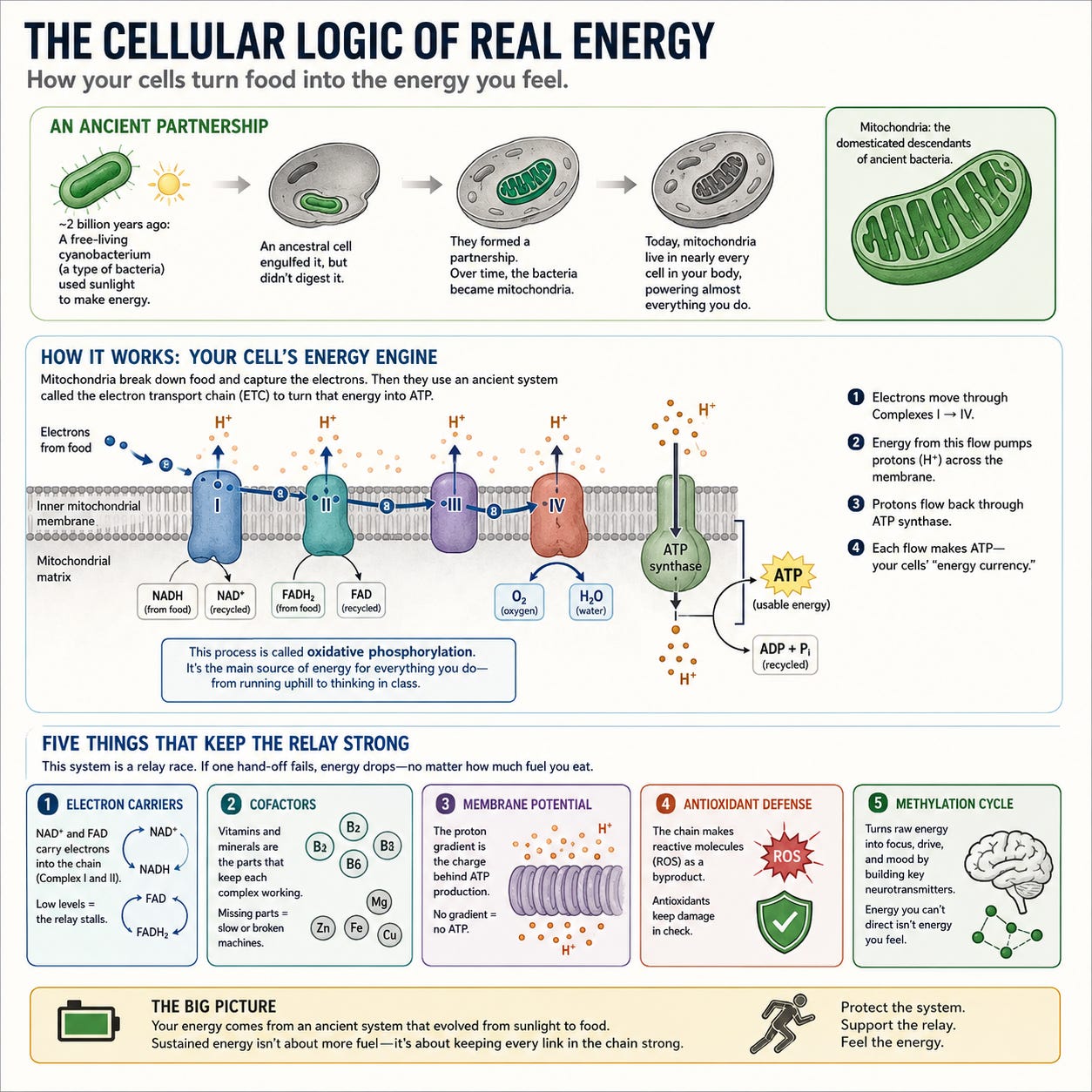
Start here, because everything else follows from it. The engine that powers almost everything you do was not, originally, yours. The mitochondria inside nearly every cell, the organelles that manufacture ATP, the currency of biological energy, are the domesticated descendants of a free-living bacterium that an ancestral cell engulfed roughly two billion years ago and never digested. That bacterium was an alphaproteobacterium, and it already knew how to do the one thing the host most needed: burn oxygen to pull energy out of food. The host kept it. The guest became an organelle. Its breathing machinery became yours.
But here is the part almost no one states correctly. The oxygen that machinery burns was not always here. For most of Earth’s history there was almost none of it. It was cyanobacteria, and for a long time cyanobacteria alone, that learned to split water with sunlight and release oxygen as the byproduct, and over hundreds of millions of years they remade the atmosphere and made aerobic life possible at all. Your mitochondria did not descend from cyanobacteria. They descended from a different bacterium entirely. But every breath those mitochondria take, every electron they pass, every molecule of ATP they build, happens inside an oxygen world that cyanobacteria created. We do not carry their lineage. We live inside their gift.
And we still meet their chemistry directly. The blue pigment in spirulina, phycocyanin, is a cyanobacterial molecule, built on the same ancient family of ring structures that forms chlorophyll in plants and heme in your blood, the two of those distinguished mostly by the metal at the center. When you take it in, you are not introducing something foreign. You are reintroducing a chemistry your cells have been reading, in one form or another, for two billion years. We will come back to it.
Here is how it works. The mitochondria take the electrons stripped from your food and pass them down a sequence of four protein complexes embedded in the inner mitochondrial membrane, the electron transport chain (ETC), using the energy released to pump protons across the membrane. The protons then flow back through an enzyme called ATP synthase, and each flow-through stamps out a fresh molecule of ATP. This process is called oxidative phosphorylation, and is a primary source of cellular energy for everything you do, from running uphill to holding a thought in working memory (Goldman Labs).
What conventional energy theory rarely says out loud is that this is a relay of electrons, and a relay is only as good as its weakest hand-off. Sustained energy is not a matter of adding fuel. It is a matter of keeping an ancient circuit intact at every link. Five things have to work:
Electron carriers must be present and abundant, NAD+ feeding Complex I, FAD feeding Complex II. These are the hands that catch and pass the electron. Deplete them and the relay stalls no matter how much fuel arrives.
The cofactors for each complex must be in place, many of them B-vitamin and trace-mineral dependent. The complexes are not abstractions. They are physical machines that seize up without their parts.
The mitochondrial membrane potential has to be intact. Without it, there is no proton gradient, and without the gradient, no ATP. This is the charge the entire ancestral design depends on.
Antioxidant defense has to keep pace with the reactive oxygen species the chain inevitably produces. Push electron flow harder and you generate more oxidative byproduct, the standard tradeoff of cellular respiration and the reason mitochondria can damage themselves with their own output.
The methylation cycle has to run cleanly, because it builds the neurotransmitters that turn raw cellular energy into felt focus and drive. Energy you cannot direct is not energy you experience.
Almost every commercial energy product ignores this entirely and reaches instead for the adrenal-sympathetic axis: caffeine, guarana, synthetic B-vitamin megadoses, quick fixes that only work for short bursts that push cortisol (which also contributes to overweight and obesity). That borrows energy from tomorrow and bills the nervous system for it. Supporting the chain itself is a different proposition. It is not stimulation. It is substrate, given to a circuit your cells have been running for two billion years.
MethylateBLUE™ was designed to feed every one of these five layers in a single two-capsule daily dose. That is the design.
The Ingredients, One by One

1. Phycocyanin — 500 mg of Blue Spirulina Extract
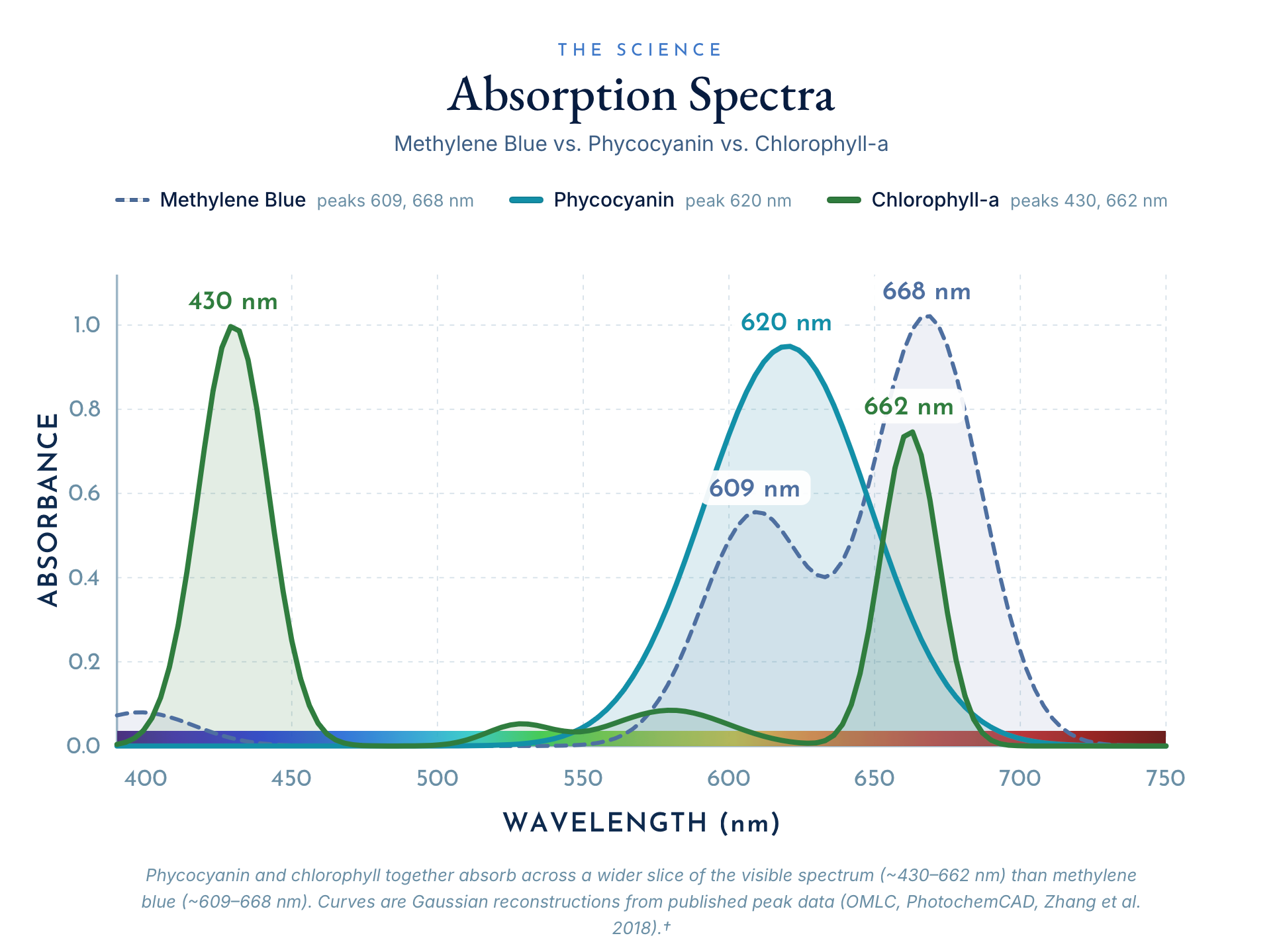
Phycocyanin is the deep-blue pigment-protein concentrated in spirulina. In photosynthetic organisms, it captures light and donates electrons. In your cells, the same chemistry has been shown across a now-extensive literature to support sustained energy. To understand why, start with the chromophore.
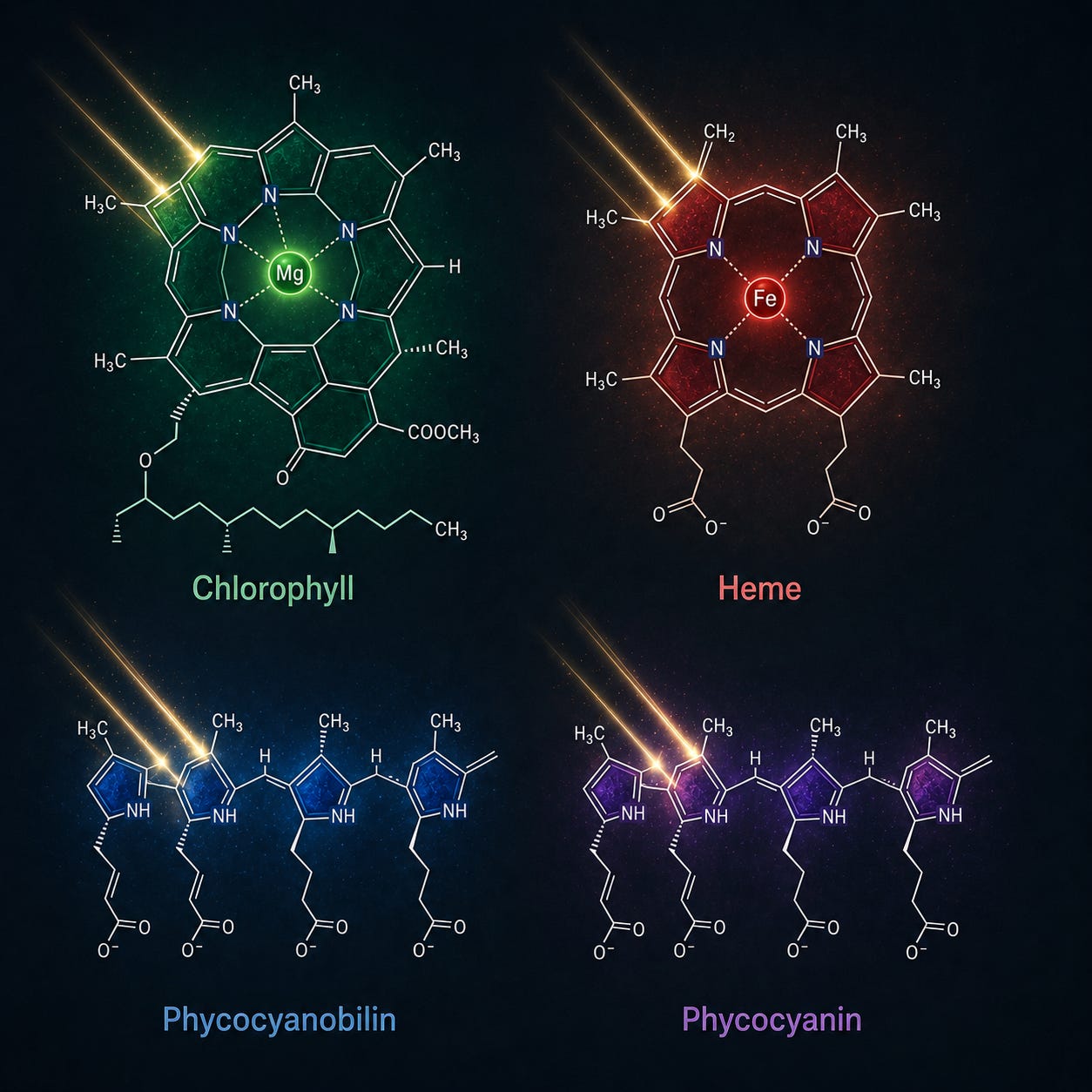
The active heart of phycocyanin is phycocyanobilin, a tetrapyrrole: four nitrogen-bearing rings, the most conserved light-and-electron-handling structure in all of biology. The same scaffold builds chlorophyll, which carries magnesium at its center, and heme, the core of your hemoglobin, which carries iron. The difference between the green pigment of plants and the red pigment of your blood is, at the center of the ring, a single swapped metal atom. Phycocyanobilin itself is nearly indistinguishable from biliverdin, the pigment your own liver produces when it breaks down heme, and from bilirubin, biliverdin’s downstream product and one of the most potent endogenous antioxidants in your bloodstream (PMC9185767 — phycocyanobilin neuropharmacology).
This is not a resemblance. It is molecular geometry, and it explains why phycocyanin behaves inside your cells less like a foreign compound and more like a signal your biology already knows how to read. Medicine exploits the same chemistry deliberately in photodynamic therapy, where a tetrapyrrole photosensitizer is introduced, light is applied, and the chromophore moves electrons to do clinical work — direct proof that these rings are not passive pigments but light-responsive electron handlers. When phycocyanobilin enters your tissue, your cells appear to recognize the family resemblance and respond accordingly. The literature documents four such responses that bear directly on energy:
Activate the Nrf2 antioxidant response. Nrf2 is the master transcription factor that switches on your body’s endogenous antioxidant enzymes — glutathione peroxidase, catalase, superoxide dismutase. A 2025 study in Zoological Research showed C-phycocyanin activates the NRF2/xCT/GPX4 pathway, enhances antioxidant activity, improves mitochondrial function, and suppresses ferroptotic cell death (PMC12940770). Tellingly, bilirubin — your own tetrapyrrole — is itself a natural Nrf2 activator, which is exactly what the structural kinship would predict. A 2016 review in Free Radical Biology & Medicine details how Nrf2 directly regulates mitochondrial biogenesis and respiration (PMC4726722).
Protect mitochondrial membrane potential. Membrane potential is the proton gradient that makes ATP synthesis possible. When phycocyanin protects that potential, every other downstream step works better.
Scavenge peroxyl and hydroxyl radicals — the same radical-quenching capacity its biliverdin-like structure would lead you to expect.
Reduce skeletal-muscle damage and fatigue. A 2024 PLOS ONE study found phycocyanin attenuates skeletal muscle damage and fatigue in exercising subjects via mitochondrial mechanisms (PLOS ONE).
In plain terms: phycocyanin protects the machinery that makes ATP while supporting the cellular pathway that builds more of it. That is the molecular definition of a non-stimulant energy supporter. I've traced the full ancestral story of this pigment — why your cells recognize it after two billion years — in We Are, In Part, Cyanobacteria.
2. Chlorophyll from Organic Spirulina — Light-Driven ATP Augmentation

The chlorophyll story is more remarkable than most readers know.
In a landmark 2014 paper in the Journal of Cell Science, Chen Xu and colleagues at Columbia University Medical Center demonstrated that mammalian mitochondria can capture light and synthesize ATP when a dietary metabolite of chlorophyll is present (PMC6518289). Isolated mouse mitochondria, brain homogenates, and live C. elegans worms all produced substantially more ATP when chlorophyll metabolites were combined with light exposure — in C. elegans, this translated to a measurable extension of lifespan.
The mechanism: chlorophyll metabolites accumulate in tissues, then catalyze the photoreduction of coenzyme Q10 — a rate-limiting step in respiration (this also creates the bioactive form of CoQ10 which known as ubiquinol in distinction from the standard form known as ubiquinone). With a chlorophyll metabolite present and red light striking the tissue, mitochondrial membrane potential rises and oxygen consumption climbs, indicating active oxidative phosphorylation.
In plain terms: chlorophyll may let your mitochondria draw a little extra energy directly from sunlight, through your skin and tissue. The early evidence is intriguing, though not yet confirmed in human clinical trials, and you can go deeper in my earlier article, We Are Beings of Light. Pair MethylateBLUE™ with sunlight or red-light therapy and you may quite literally be adding photons to the ATP equation. This would be energy you can’t borrow back; it arrives from entirely outside the body’s stored reserves, and the same mechanism appears to enhance the efficiency of your own Krebs cycle.
3. Fermented Nicotinamide — The NAD+ Precursor at Complex I
NAD+ — nicotinamide adenine dinucleotide — is the most important coenzyme you have never heard of. Every cell in the body uses it (NIH/Molecular Metabolism review on NAD+).
Here is what NAD+ does:
It is the primary electron carrier that feeds Complex I of the electron transport chain. When food is broken down via glycolysis, beta-oxidation, and the TCA cycle, the electrons are loaded onto NAD+, which becomes NADH. NADH then delivers those electrons to Complex I to start the cascade that makes ATP (Wikipedia — NAD; Goldman Labs). One NADH yields roughly three ATP molecules.
It is the substrate that powers sirtuins (longevity-associated deacetylases), PARPs (DNA repair enzymes), and CD38 (immune signaling).
It declines with age. NAD+ levels drop measurably after 30, and that decline is among the most-cited cellular mechanisms behind biological aging.
Fermented nicotinamide has higher bioavailability and is a direct precursor to NAD+. Inside the cell, it enters the salvage pathway — the dominant route by which mammals make NAD+, accounting for ~85% of total NAD+ production (PMC7973386). Unlike higher-dose niacin, nicotinamide does not cause the characteristic flush. Unlike NMN or NR (the trendier NAD+ precursors), it has a long history of safe use, an unambiguous biochemical fate, and a price point that lets it be dosed properly.
When you feed your cells more NAD+ raw material, Complex I has more electron carriers to receive from. That is not a metaphor. It is the literal substrate-availability step in the chain that generates the vast majority of your daily ATP. And this is why you feel more, and better “crash free” energy.
The Four Methylated B Vitamins — Cofactors at Every Step of Energy Metabolism
B-vitamin deficiency, even sub-clinical, directly impairs mitochondrial metabolism of glucose, fats, and amino acids through the citric acid cycle and electron transport chain (Permanente Journal — B Vitamins in Medicine). That is not a wellness claim. It is fundamental biochemistry.
This is not theoretical for me. I have taken a whole-food B-complex nearly every day for the better part of twenty years, from companies I still respect like Megafood, Garden of Life, and New Chapter, because I understood early that these cofactors are not optional. MethylateBLUE is the formula I finally built when I wanted every one of them in its active, methylated form, in one place, alongside the pigment chemistry I'd spent years writing about. In the most literal sense, it's the second supplement I made for myself first.
MethylateBLUE™ uses the four B vitamins in their active, bioavailable, methylated forms — the same forms the body uses internally. About 40% of the population carries an MTHFR variant that limits their ability to convert standard folic acid and cyanocobalamin into anything useful; the methylated forms bypass that bottleneck entirely.
Riboflavin as R-5-Phosphate (1.95 mg). Riboflavin (vitamin B2) is the precursor to FAD and FMN, the flavin coenzymes that power Complex I and Complex II of the electron transport chain alongside NAD+ (PMC7973386; ScienceDirect — Vitamin B Complex). Without functional FAD, Complex II grinds to a halt. R-5-Phosphate is the form your cells use directly — no conversion step.
Niacin as Nicotinamide (44 mg). Already covered above — the precursor to NAD+ for Complex I.
Vitamin B6 as Pyridoxal 5’-Phosphate (2.55 mg). P-5-P is the active form of B6 and is a cofactor for over 140 enzymatic reactions. Among its most important roles: synthesis of dopamine, serotonin, norepinephrine, and GABA — the neurotransmitters that turn cellular energy into focused attention and motivated drive.
Folate as L-5-MTHF (400 mcg DFE) + B12 as Methylcobalamin (12 mcg). These two work together to run the methylation cycle — about a billion methylation reactions per second, including the synthesis of those same neurotransmitters and the conversion of homocysteine back to methionine. B12 also enables the conversion of L-methylmalonyl-CoA to succinyl-CoA, which directly enters the citric acid cycle and plays “an important role in the production of energy from lipids and proteins” (Linus Pauling Institute — Vitamin B12).
In other words: the B vitamins in MethylateBLUE™ feed both the energy production line and the neurotransmitter line. That dual role is why people describe the felt experience as “calm, lit-up focus” rather than the wired-but-jittery state caffeine produces — the latter has only the neurotransmitter half, with no cellular energy to back it.
5. The Nine Peptide-Powered Minerals™ — Mitochondrial Cofactors at Intracellular Doses

A central insight of mitochondrial biology is that the electron transport chain and the antioxidant enzyme system are mineral-dependent. Without the right trace metals in the right intracellular compartments, the whole apparatus fails.
MethylateBLUE™ delivers nine essential minerals, every one of them bound to the di- and tri-peptides naturally present in organic spirulina (the VerdeMins™ process), which is the form your gut has evolved a transporter for over two million years.
Magnesium (110 mg). Magnesium is arguably the most under-appreciated energy cofactor in the body. It is required for F0/F1-ATP synthase — the terminal enzyme of oxidative phosphorylation, the molecular motor that actually stamps out ATP (PMC5516748 — Mg2+ and OXPHOS). It activates OGDH in the citric acid cycle. And a 2023 Nature Communications Biology study showed intracellular magnesium directly protects mitochondria from oxidative stress damage (nature.com/s42003-023-05247-6). Every molecule of ATP in your body exists as Mg-ATP — magnesium and ATP travel together.
Iron (9 mg). Iron is at the catalytic center of the cytochromes inside Complexes I, II, III, and IV of the electron transport chain. No iron, no electron flow, no ATP.
Iodine (75 mcg) + Selenium (55 mcg). This is the thyroid pair. Iodine is the structural backbone of T4 and T3 — the thyroid hormones that set your basal metabolic rate (StatPearls — Thyroid Hormone; Physiological Reviews — TH Metabolism). Selenium is the cofactor for the deiodinase enzymes that convert inactive T4 into active T3 inside target tissues. T3 then increases gene expression of Na+/K+ ATPase, increases oxygen consumption, and raises body temperature. If you feel persistently cold and persistently tired, your iodine-selenium axis is one of the first places to look.
Zinc (10 mg), Copper (0.9 mg), Manganese (2.3 mg). This is the SOD enzyme triad — Cu/Zn-SOD in the cytoplasm and Mn-SOD inside the mitochondrial matrix. These are your body’s first-line antioxidant defenses against the reactive oxygen species that ETC activity generates. Without them, mitochondrial damage accumulates and ATP output declines.
Chromium (35 mcg). Chromium enhances insulin sensitivity and helps maintain steady blood glucose (Linus Pauling Institute — Chromium; Journal of Nutritional Biochemistry on chromium and insulin signaling). A randomized clinical trial in Diabetes Care showed chromium picolinate improved glycemic control and insulin sensitivity in type 2 diabetic subjects (Diabetes Journals — Chromium Picolinate Trial). For sustained energy, steady glucose is the foundation under everything else.
Molybdenum (45 mcg). Cofactor for sulfite oxidase, xanthine oxidase, and aldehyde oxidase — required for sulfur metabolism and detox.
Notice the pattern. Every mineral here either lives inside the electron transport chain, powers an antioxidant enzyme that protects it, or regulates the hormonal signals that set its tempo. Nothing is decorative.
Part III — Contrast: What Sugar, Coffee, Tea, Alcohol, and Stimulants Actually Do
Each of these substances “works” — meaning users feel a real subjective lift — but the cellular mechanism is completely different from the one MethylateBLUE™ engages. Understanding the difference is the entire point.
Sugar (and refined carbohydrates) — The Blood-Glucose Spike-and-Crash
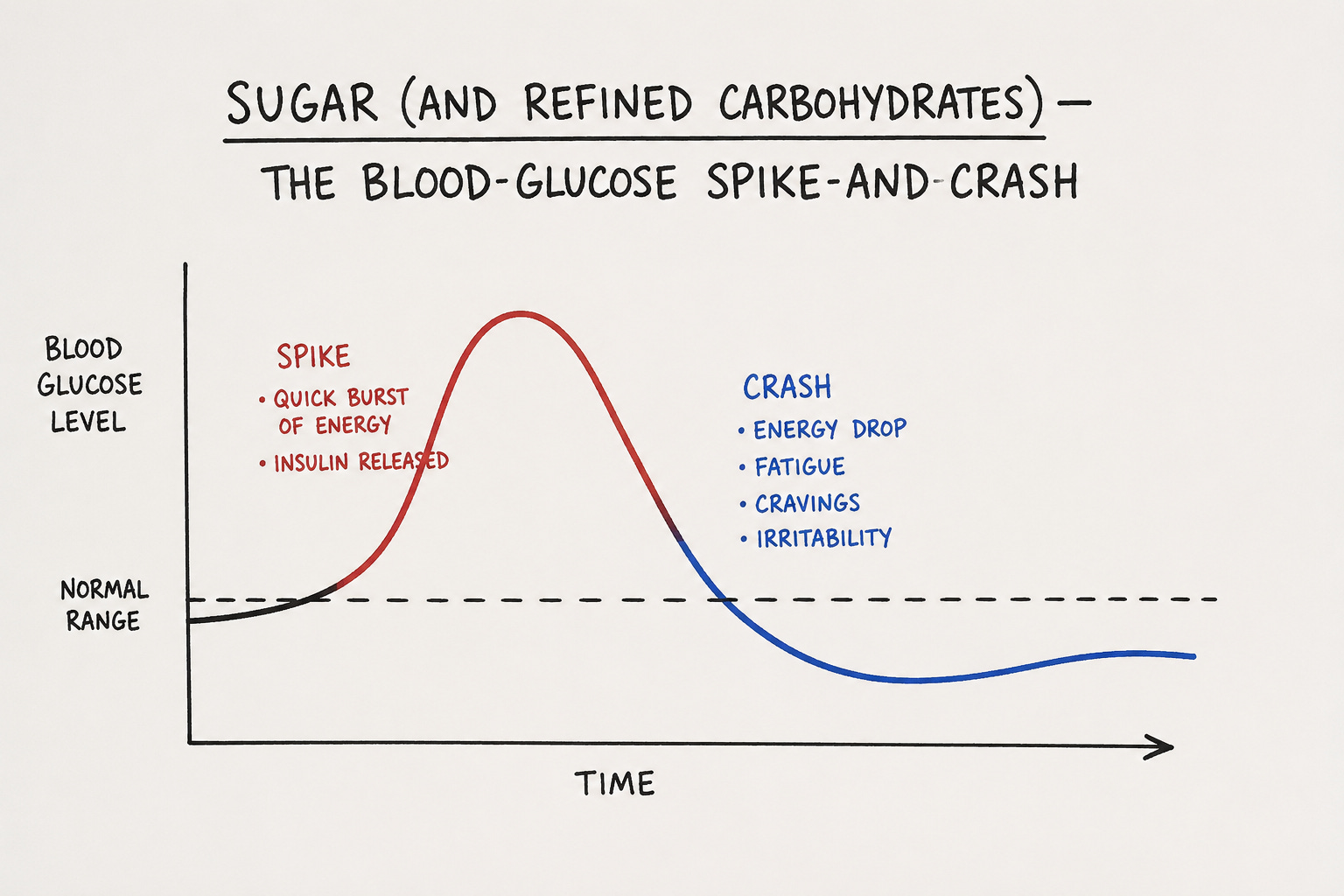
Refined sugar floods the bloodstream with glucose faster than your insulin response can match. The pancreas overshoots, secretes more insulin than the meal actually required, and that excess insulin then drives blood glucose below the pre-meal baseline. The result is the classic reactive hypoglycemia crash: shakiness, irritability, brain fog, sudden hunger (Mayo Clinic; Wikipedia — Reactive Hypoglycemia; Şişli Etfal Tıp Bülteni).
You did not gain energy. You took out a glucose loan, paid it back with insulin interest, and ended up below where you started. Repeated over years, this drives insulin resistance — the upstream condition behind type 2 diabetes.
MethylateBLUE™ does the opposite at this exact node: chromium supports insulin sensitivity, magnesium supports insulin signaling, and the rest of the formula supports the mitochondria that actually burn the glucose your meal provides.
Coffee and Tea (Caffeine) — Blocking the Tired Signal, Not Adding Energy
This one is worth quoting directly because it surprises most people:
“Caffeine does not provide energy — it acts by blocking adenosine receptors in the brain. The post-caffeine ‘crash’ results from accumulated adenosine binding once caffeine dissociates.” (Ultrahuman)
Adenosine is the molecule your brain uses to signal sleep pressure. As neurons work through the day, adenosine accumulates and binds to A1 and A2A receptors, slowing neural activity to encourage rest. Caffeine has a molecular shape similar enough to adenosine that it occupies those receptors without activating them (Life Sciences — adenosine receptor antagonists).
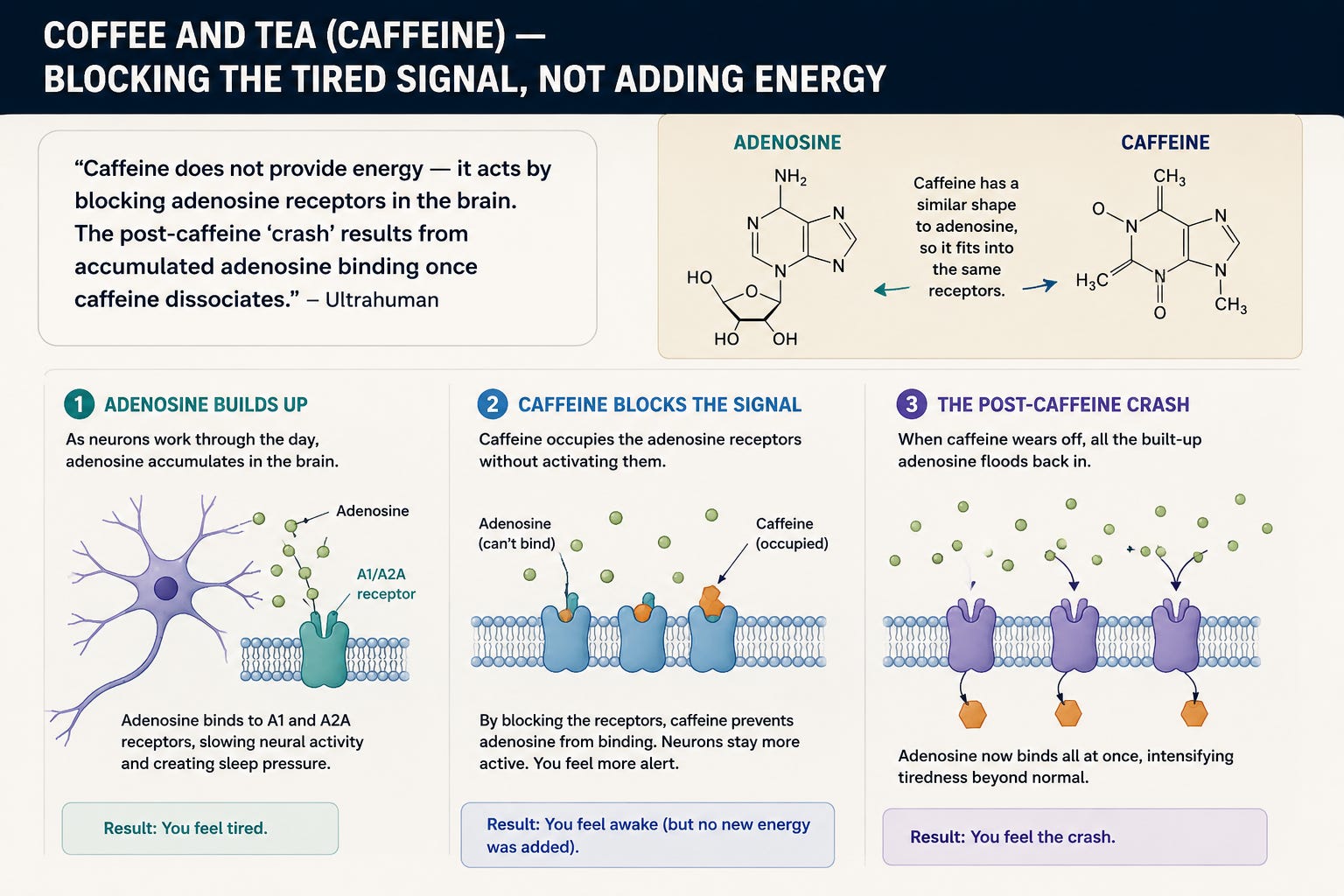
This produces three problems:
It is a perception trick, not real energy. Adenosine keeps accumulating in the background; caffeine just hides the signal.
The crash is built into the mechanism. When caffeine dissociates, all that accumulated adenosine — now at higher concentrations than it would normally have reached — binds at once. Fatigue arrives in a sharp spike rather than a gentle slope (Ultrahuman).
Tolerance is enforced biology. With chronic intake, the brain upregulates adenosine receptors to compensate for the constant blockade. You need more caffeine to produce the same effect, and your baseline focus erodes (Ultrahuman).
Caffeine is a fine acute tool. As a daily energy strategy, it is a debt instrument with a built-in repayment schedule. MethylateBLUE™ supports the cellular machinery underneath, so the perception of energy reflects the reality.
Alcohol — Direct Mitochondrial Damage
Alcohol is the only item on this list that literally damages the energy-producing organelles.
A 2022 Autophagy study from MUSC demonstrated that both acute and chronic ethanol exposure causes mitochondrial depolarization — collapse of the membrane potential that ATP synthesis depends on (MUSC News; Gastroenterology review on alcohol and mitochondria). Damaged mitochondria then get cleared via mitophagy. Acute alcohol exposure can be repaired by replication of healthy mitochondria; chronic exposure leads to net mitochondrial depletion, inflammation, fibrosis, and ultimately liver disease.
Alcohol also consumes massive quantities of NAD+ during its metabolism (Frontiers in Immunology — NAD+ depletion and mitochondrial dysfunction). Less NAD+ means less ATP. This is part of why hangover fatigue is so severe — you have actively drained the electron carrier the entire chain depends on.
MethylateBLUE™ feeds NAD+ directly via fermented nicotinamide. It protects mitochondrial membrane potential via phycocyanin. It is, in mechanistic terms, the architectural opposite of alcohol’s effect on the cell.
Nicotine — Receptor Hijack and Built-In Tolerance

Billions of servings of nicotine-containing products enter human bodies around the world, each day. Even though these products are clearly labeled addictive and harmful, but why?
The reason has to do with simple biochemistry.
Nicotine binds and activates nicotinic acetylcholine receptors (nAChRs) in the brain, producing acute releases of dopamine, norepinephrine, and acetylcholine — hence the lift, the focus, and the calm-but-alert feeling. The trap is in the cellular response: chronic nicotine exposure causes those same receptors to upregulate — the brain manufactures more of them to compensate (PMC2728164 — nicotinic receptor upregulation; PMC4780670 — nAChR maturation and assembly). The result is escalating dose for diminishing effect, the textbook trajectory of dependence.
Notice the family resemblance to caffeine: both work by manipulating receptor signaling without delivering any cellular substrate, both build tolerance through receptor adaptation, both require escalation to maintain effect.
MethylateBLUE™ does not bind receptors, simulating neurotransmitter activity. It feeds substrates. There is no downregulation pathway for “having enough magnesium” or “having abundant NAD+.” Your cells use what’s there and the rest is excreted.
Energy Drinks — All of the Above in One Can
Caffeine + sugar + sometimes nicotine analogues + occasionally taurine and B-vitamins as a fig leaf. The B-vitamins in most energy drinks are the cheap unmethylated forms (cyanocobalamin and folic acid), which the ~40% of the population with MTHFR variants poorly convert (explainer in the MethylateBLUE FAQ). You get the receptor manipulation of caffeine, the glucose spike-crash of sugar, and a label that mentions vitamins without supplying their bioactive forms.
Part IV — The Architectural Difference, Summarized
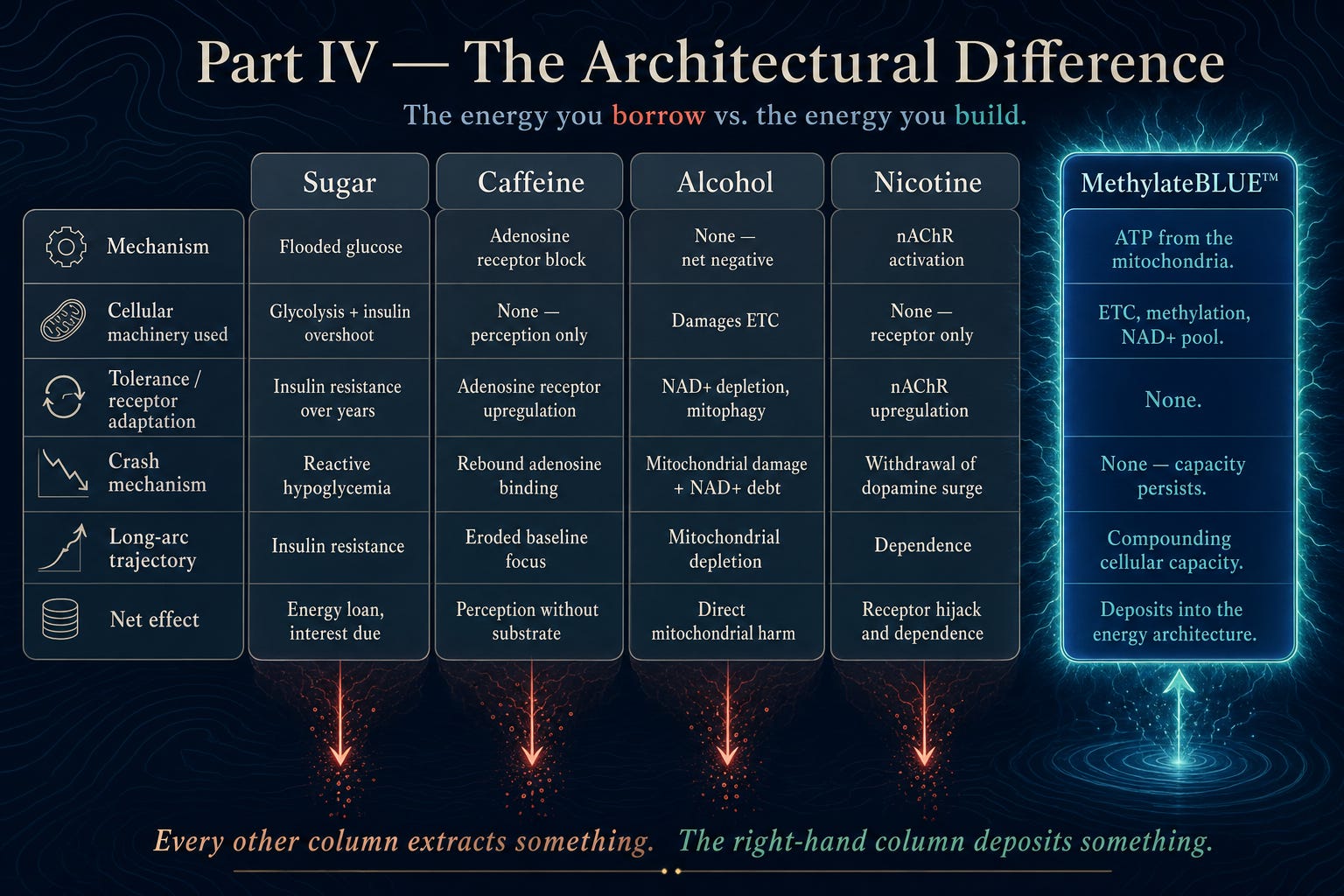
The graphic above compresses the entire argument of this section into a single frame. Each column represents a familiar “energy” input—sugar, caffeine, alcohol, nicotine—and the MethylateBLUE™ column on the right. Row by row, it shows not just how they feel in the moment, but which cellular machinery they recruit, what kind of tolerance they induce, how they crash, and where they leave you over the long arc.
In the first four columns, the pattern is the same. Sugar floods the bloodstream with glucose and drives reactive hypoglycemia. Caffeine blocks adenosine receptors without adding any ATP. Alcohol directly damages mitochondria and drains NAD+. Nicotine hijacks nicotinic receptors and builds dependence. Every one of these pathways is fundamentally extractive: they manipulate signals, deplete cofactors, and erode capacity over time.
The MethylateBLUE™ column is architected in the opposite direction. Its “mechanism” row points to ATP coming from the mitochondria themselves, supported by intact electron transport, methylation, and an expanded NAD+ pool. Instead of tolerance, there is no receptor adaptation. Instead of a crash, capacity persists. Instead of a trajectory toward insulin resistance, depleted focus, mitochondrial loss, or dependence, the long‑arc row ends in “compounding cellular capacity.”
Visually, the muted, downward‑tilted arrows under sugar, caffeine, alcohol, and nicotine mark the quiet drain they impose on the system, while the numinous electrical halo around MethylateBLUE™ signals a different architecture: one that deposits into the energy‑producing organelles instead of borrowing from them. In a glance, the graphic is meant to make the main claim of this article unmistakable: all the usual “energy” tools extract; this formula is designed to build.
Part V — The Compound-Interest Effect
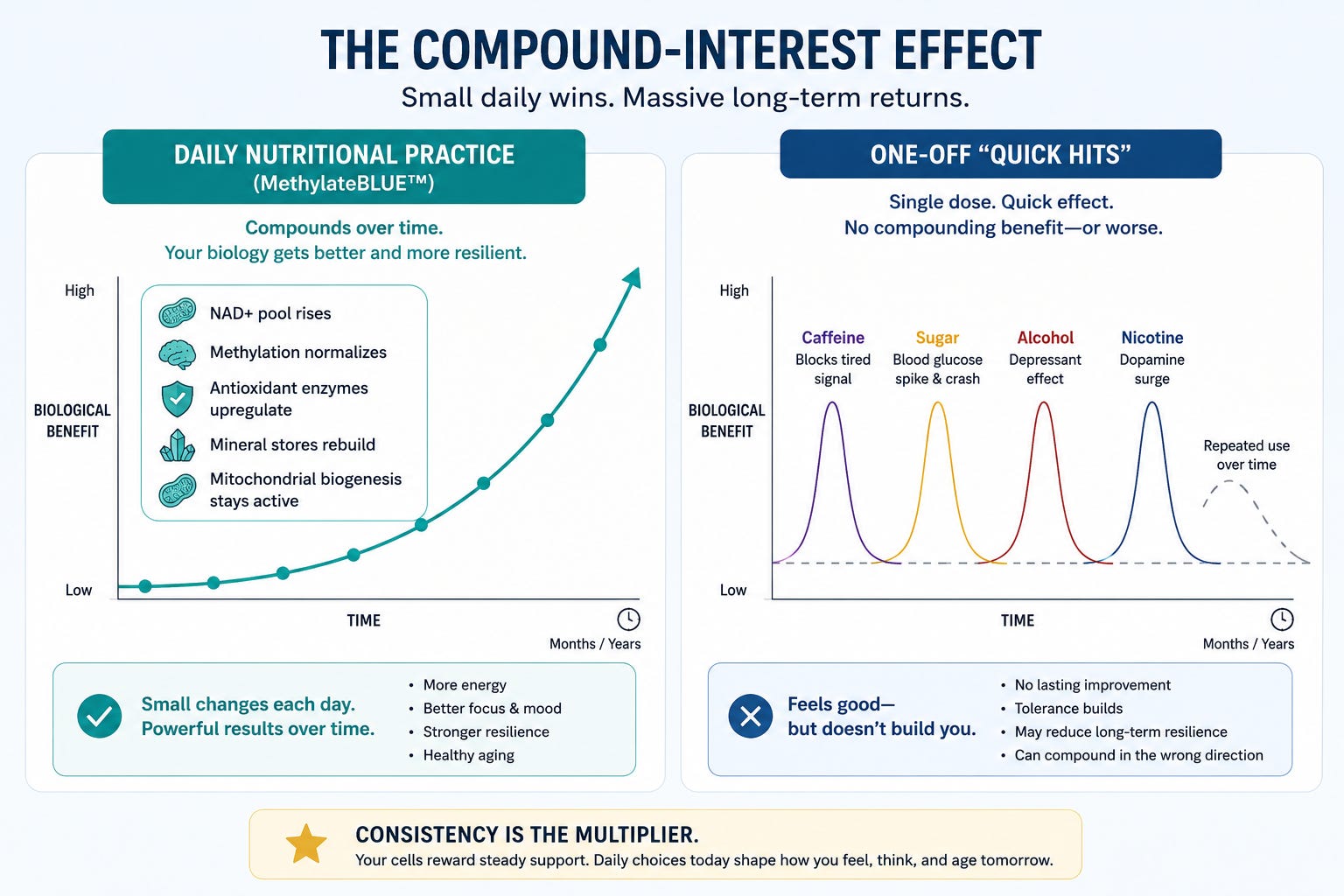
There is one more concept worth naming, because it is the practical, lived-experience reason that a daily nutritional practice eventually outperforms any single-hit intervention — including IV drips, single-day megadoses, and one-off “biohacks.”
A single dose of a nutrient delivers a single dose of effect. A daily practice, sustained over months and years, compounds. Your NAD+ pool gradually rises. Your methylation status normalizes. Your antioxidant enzymes upregulate to match your antioxidant intake. Your mineral stores rebuild. Your mitochondrial biogenesis pathways stay activated. Each of those changes is small on any given day and large after a year.
This is why the MethylateBLUE™ product page describes a felt effect within hours, and a build of mitochondrial capacity, antioxidant defense, and methylation status over 8 to 12 weeks of consistent use. The first arrives because B-vitamin and NAD+ pathways respond quickly. The second arrives because cellular biology rewards consistency.
By contrast, every substance on the comparison list above either delivers no compounding benefit at all (caffeine, nicotine) or compounds in the wrong direction (sugar, alcohol). The longer these quick‑hit inputs are used as a primary strategy, the more research suggests they may be associated with less efficient metabolic and neural responses over time—even if the subjective ‘hit’ feels similar.
Part VI — What This Looks Like in Practice
Two capsules with breakfast. Once a day. Six to eight weeks before you fairly judge the full effect, though many people feel a clean, non-stimulating lift on day one.
What you are doing, over those weeks, is:
Saturating your NAD+ pool from the fermented nicotinamide.
Bypassing any MTHFR conversion bottleneck with the four already-methylated B vitamins.
Building intracellular concentrations of nine essential minerals via the peptide-bound delivery system.
Activating your endogenous antioxidant enzymes via phycocyanin and Nrf2.
Protecting mitochondrial membrane potential.
Supplying the chlorophyll metabolite that allows your mitochondria to convert sunlight into supplemental ATP when you spend time outdoors.
You are, in short, building the body that produces sustained, focused energy from within — instead of stimulating the body you already have into spending tomorrow’s reserves today.
I did not set out to make a supplement. I set out to understand why energy that lasts feels so different from energy that spikes, and the answer led me back two billion years into the oldest machinery we carry. The product is what followed. But the understanding came first, and it's the part I most want you to keep, whether or not you ever try the formula.
Ancient Pigment. Modern Biochemistry.
That is the formula. That is the philosophy. That is the difference.
Taken as part of a broader lifestyle that includes movement, a nutrient‑dense diet, and adequate sleep, MethylateBLUE™ is meant to support the body’s own architecture for sustained, focused energy, rather than replace medical care or treat any condition

Sources Cited
Goldman Labs, “What is Nicotinamide Adenine Dinucleotide?” (goldmanlaboratories.com)
Molecular Metabolism, “Role of NAD+ in regulating cellular and metabolic signaling pathways” (PMC7973386)
Wikipedia, “Nicotinamide adenine dinucleotide” (wikipedia.org)
Zoological Research, “Rescuing fertility: C-Phycocyanin prevents ovarian damage via NRF2/xCT/GPX4” (PMC12940770)
Free Radical Biology & Medicine, “The emerging role of Nrf2 in mitochondrial function” (PMC4726722)
Current Neuropharmacology, “C-Phycocyanin-derived Phycocyanobilin as a Potential Neuroprotective Agent” (PMC9185767)
PLOS ONE, “Phycocyanin attenuates skeletal muscle damage and fatigue” (journals.plos.org)
Xu et al., Journal of Cell Science, “Light-harvesting chlorophyll pigments enable mammalian mitochondria to capture photonic energy and produce ATP” (PMC6518289)
The Permanente Journal, “B Vitamins: Functions and Uses in Medicine” (PMC9662251)
Nutrients, “B Vitamins and the Brain: Mechanisms, Dose and Efficacy” (PMC4772032)
Linus Pauling Institute, “Vitamin B12” (lpi.oregonstate.edu)
Linus Pauling Institute, “Chromium” (lpi.oregonstate.edu)
Oxidative Medicine and Cellular Longevity, “The Involvement of Mg2+ in Regulation of Cellular and Mitochondrial Function” (PMC5516748)
Nature Communications Biology, “Intracellular Mg2+ protects mitochondria from oxidative stress” (nature.com)
StatPearls, “Physiology, Thyroid Hormone” (NCBI Bookshelf)
Physiological Reviews, “Thyroid Hormone Regulation of Metabolism” (PMC4044302)
Diabetes Care, “Chromium Picolinate Supplementation Attenuates Body Weight Gain” (Diabetes Journals)
Journal of Nutritional Biochemistry, “Molecular Mechanisms of Chromium in Alleviating Insulin Resistance” (PMC3308119)
Mayo Clinic, “Reactive hypoglycemia” (mayoclinic.org)
Şişli Etfal Hastanesi Tıp Bülteni, “Postprandial Reactive Hypoglycemia” (PMC7192270)
Wikipedia, “Reactive hypoglycemia” (wikipedia.org)
Ultrahuman, “Caffeine and adenosine: The reason behind the coffee crash” (blog.ultrahuman.com)
Life Sciences, “Using caffeine and other adenosine receptor antagonists and agonists as therapeutic tools” (PMC4115368)
MUSC News, “Excessive degradation of mitochondria is the tipping point” (musc.edu)
Gastroenterology, “Alcohol and Mitochondria: A Dysfunctional Relationship” (PMC1868435)
Frontiers in Immunology, “Epithelial NAD+ depletion drives mitochondrial dysfunction” (PMC10512956)
Biochemical Pharmacology, “Nicotine-induced Upregulation of Nicotinic Receptors” (PMC2728164)
Genes, Brain and Behavior, “Nicotinic acetylcholine receptors: upregulation, age-related effects” (PMC4780670)
†These statements have not been evaluated by the Food and Drug Administration. MethylateBLUE™ is not intended to diagnose, treat, cure, or prevent any disease.
No hay comentarios:
Publicar un comentario